Игра эволюция рнк и рождение информации. В чем заключается теория РНК-мира? Абиогенный синтез РНК
Мир РНК - гипотетический этап возникновения жизни на Земле, когда как функцию хранения генетической информации, так и катализ химических реакций выполняли ансамбли молекул рибонуклеиновых кислот. Впоследствии из их ассоциаций возникла современная ДНК-РНК-белковая жизнь, обособленная мембраной от внешней среды. Идея мира РНК была впервые высказана Карлом Вёзе в 1968 году, позже развита Лесли Орджелом и окончательно сформулирована Уолтером Гильбертом в 1986 году.
Краткое изложение
В живых организмах практически все процессы происходят в основном благодаря ферментам белковой природы. Белки, однако, не могут самореплицироваться и синтезируются в клетке de novo на основании информации, заложенной в ДНК. Но и удвоение ДНК происходит только благодаря участию белков и РНК. Образуется замкнутый круг, из-за которого, в рамках теории самозарождения жизни приходилось признать необходимость не только абиогенного синтеза обоих классов молекул, но и спонтанного возникновения сложной системы их взаимосвязи.
В начале 1980-х годов в лаборатории Т. Чека и С. Олтмана в США была открыта каталитическая способность РНК. По аналогии с ферментами РНК-катализаторы были названы рибозимами, за их открытие Томасу Чеку в 1989 году была присуждена Нобелевская премия по химии. Более того, оказалось, что активный центр рибосом содержит большое количество рРНК. Также РНК способны создавать двойную цепочку и самореплицироваться.
Таким образом, РНК могли существовать полностью автономно, катализируя «метаболические» реакции, например, синтеза новых рибонуклеотидов и самовоспроизводясь, сохраняя из «поколения» в «поколение» каталитические свойства. Накопление случайных мутаций привело к появлению РНК, катализирующих синтез определённых белков, являющихся более эффективным катализатором, в связи с чем эти мутации закреплялись в ходе естественного отбора. С другой стороны возникли специализированные хранилища генетической информации - ДНК. РНК сохранилась между ними как посредник.
Роль РНК в современном мире
Следы мира РНК остались в современных живых клетках, причём РНК участвует в критически важных процессах жизнедеятельности клетки:
1) Основной носитель энергии в клетках - АТФ - это рибонуклеотид, а не дезоксирибонуклеотид.
2) Биосинтез белка почти целиком осуществляется с помощью различных видов РНК:
· матричные РНК являются матрицей для синтеза белка в рибосомах;
· транспортные РНК доставляют аминокислоты к рибосомам и реализуют генетический код;
· рибосомная РНК составляет активный центр рибосом, катализирующий образование пептидной связи между аминокислотами.
3) Для репликации ДНК также критически важна РНК:
· для начала процесса удвоения ДНК необходима РНК-«затравка» (праймер);
· для бесконечного удвоения ДНК, не ограниченного пределом Хейфлика, в эукариотических клетках производится постоянное восстановление концевых участков хромосом (теломер) ферментом теломеразой, в состав которого входит РНК-матрица.
4) В процессе обратной транскрипции информация из РНК переписывается в ДНК.
5) В процессе созревания РНК используются различные РНК, не кодирующие белки, включая малые ядерные РНК, малые ядрышковые РНК.
Кроме того, многие вирусы хранят свой генетический материал в виде РНК и поставляют в заражённую клетку РНК-зависимую РНК-полимеразу для его репликации.
Абиогенный синтез РНК
Абиогенный синтез РНК из более простых соединений не продемонстрирован экспериментально в полной мере. В 1975 году Манфред Сампер и Рудигер Льюс в лаборатории Эйгена продемонстрировали, что в смеси, вообще не содержащей РНК, а содержащей только нуклеотиды и Qβ-репликазу, может при определённых условиях спонтанно возникнуть самореплицирующаяся РНК.
В 2009 году группе учёных из университета Манчестера под руководством Джона Сазерленда удалось продемонстрировать возможность синтеза уридина и цитидина с высокой эффективностью и степенью закрепления результата реакции (а также с возможностью накопления конечных продуктов) в условиях ранней Земли. В то же время, хотя абиогенный синтез пуриновых оснований продемонстрирован достаточно давно (в частности, аденин является пентамером синильной кислоты), их гликозилирование свободной рибозой аденозина и гуанозина пока показано лишь в малоэффективном варианте.
Эволюция РНК
Способность молекул РНК к эволюции была наглядно продемонстрирована в ряде экспериментов. Ещё до открытия каталитической активности РНК такие эксперименты проводили Лесли Оргел с коллегами в Калифорнии. Они добавляли к пробирке с РНК яд – бромид этидия, ингибирующий синтез РНК. Сначала темп синтеза был замедлен ядом, но примерно после девяти «пробирочных поколений» эволюции в процессе естественного отбора вывелась новая порода РНК, стойкая к яду. Путём последовательного удвоения доз яда была выведена порода РНК, стойкая к очень высоким его концентрациям. Всего в эксперименте сменилось 100 пробирочных поколений (и намного больше поколений РНК, т.к. поколения сменялись и внутри каждой пробирки). Хотя в этом эксперименте РНК-репликаза добавлялась в раствор самими экспериментаторами, Оргел обнаружил, что РНК способны и к спонтанному самокопированию, без добавления фермента, правда, намного медленнее.
Дополнительный эксперимент был позже проведён в лаборатории немецкой школы Манфреда Ейгена. Он обнаружил спонтанное самозарождение молекулы РНК в пробирке с субстратом и РНК-репликазой. Она была создана постепенно нарастающей эволюцией.
После открытия каталитической активности РНК (рибозимов) их эволюция в автоматизированном устройстве под управлением компьютера наблюдалась в экспериментах Брайана Пегеля и Джеральда Джойса из Исследовательского института имени Скриппса в Калифорнии в 2008 году. Фактором, играющим роль давления отбора, являлась ограниченность субстрата, куда входили олигонуклеотиды, которые рибозим распознавал и присоединял к себе, и нуклеотиды для синтеза РНК и ДНК. При построении копий иногда случались дефекты - мутации - влияющие на их каталитическую активность (для ускорения процесса несколько раз смесь подвергалась мутированию с помощью полимеразной цепной реакции с использованием "неточных" полимераз). По этому признаку и происходил отбор молекул: наиболее быстро копирующиеся молекулы быстро начинали доминировать в среде. Затем 90% смеси удалялось, а вместо этого добавлялась свежая смесь с субстратом и ферментами, и цикл повторялся снова. За 3 суток каталитическая активность молекул за счёт всего 11 мутаций увеличилась в 90 раз.
Эти эксперименты доказывают, что первым молекулам РНК не нужно было обладать достаточно хорошими каталитическими свойствами. Они развились потом в ходе эволюции под действием естественного отбора.
В 2009 году канадские биохимики из Монреальского университета К. Боков и С. Штейнберг, изучив основную составляющую рибосомы бактерии Escherichia coli, молекулу 23S-рРНК, показали, каким образом из относительно небольших и простых рибозимов мог развиться механизм белкового синтеза. Молекула была подразделена на 60 относительно самостоятельных структурных блоков, основным из которых является каталитический центр (пептидил-трансферазный центр, PTC, peptidyl-transferase centre), ответственный за транспептидацию (образование пептидной связи). Было показано, что все эти блоки можно последовательно отсоединять от молекулы без разрушения её оставшейся части до тех пор, пока не останется один лишь транспептидационный центр. При этом он сохраняет способность катализировать транспептидацию. Если каждую связь между блоками молекулы представить в виде стрелки, направленной от того блока, который при отрыве не разрушается, к тому блоку, который разрушается, то такие стрелки не образуют ни одного замкнутого кольца. Если бы направление связей было случайным, вероятность этого составляла бы менее одной миллиардной. Следовательно, такой характер связей отражает последовательность постепенного добавления блоков в процессе эволюции молекулы, который исследователям удалось детально реконструировать. Таким образом, у истоков жизни мог стоять сравнительно простой рибозим - PTC-центр молекулы 23S-рРНК, к которому затем добавлялись новые блоки, совершенствуя процесс синтеза белка. Сам PTC состоит из двух симметричных лопастей, каждая из которых удерживает CCA"-хвостик одной молекулы тРНК. Предполагается, что такая структура возникла в результате дупликации (удвоения) одной исходной лопасти. Методом искусственной эволюции были получены функциональные РНК (рибозимы), способные катализировать транспептидацию. Структура этих искусственно выведенных рибозимов очень близка к структуре той проторибосомы, которую «вычислили» авторы.
Свойства объектов мира РНК
О том, как выглядели самовоспроизводящиеся РНК системы, есть разные предположения. Чаще всего постулируется необходимость агрегирующих РНК мембран или размещения РНК на поверхности минералов и в поровом пространстве рыхлых пород. В 1990-е годы А. Б. Четвериным с сотрудниками была показана способность РНК формировать молекулярные колонии на гелях и твёрдых субстратах при создании им условий для репликации. Происходил свободный обмен молекулами, которые при столкновении могли обмениваться участками, что показано экспериментально. Вся совокупность колоний в связи с этим быстро эволюционировала.
После возникновения белкового синтеза колонии, умеющие создавать ферменты, развивались успешнее. Ещё более успешными стали колонии, сформировавшие более надёжный механизм хранения информации в ДНК и, наконец, отделившиеся от внешнего мира липидной мембраной, препятствующей рассеиванию своих молекул.
Пре-РНК миры
Биохимик Р. Шапиро критикует гипотезу РНК-мира, считая, что вероятность спонтанного возникновения РНК, обладающей каталитическими свойствами, очень низка. Взамен гипотезы «вначале была РНК», он предлагает гипотезу «вначале был метаболизм», то есть возникновение комплексов химических реакций - аналогов метаболических циклов - с участием низкомолекулярных соединений, протекающих внутри компартментов - пространственно ограниченных самопроизвольно образовавшимися мембранами или иными границами раздела фаз - областей. Эта концепция близка к коацерватной гипотезе абиогенеза, предложенной А. И. Опариным в 1924 году.
Другой гипотезой абиогенного синтеза РНК, призванной решить проблему низкой оценочной вероятности синтеза РНК, является гипотеза мира полиароматических углеводородов, предложенная в 2004 году и предполагающая синтез молекул РНК на основе стека из полиароматических колец.
Фактически, обе гипотезы «пре-РНК миров» не отвергают гипотезу мира РНК, а модифицируют её, постулируя первоначальный синтез реплицирующихся макромолекул РНК в первичных метаболических компартментах, либо на поверхности ассоциатов, отодвигая «мир РНК» на вторую стадию абиогенеза.
Академик РАН А. С. Спирин считает, что РНК-мир не мог появиться и существовать на Земле, и рассматривает вариант внеземного (в первую очередь на кометах) происхождения и эволюции РНК-мира.
© А.С. Спирин
БИОСИНТЕЗ БЕЛКОВ, МИР РНКИ ПРОИСХОЖДЕНИЕ ЖИЗНИ
А.С. Спирин
Спирин Александр Сергеевич - академик, директор Института белка РАН, член Президиума РАН.
Почти полвека тому назад, в 1953 г., Д. Уотсон и Ф. Крик открыли принцип структурной (молекулярной) организации генного вещества - дезоксирибонуклеиновой кислоты (ДНК) . Структура ДНК дала ключ к механизму точного воспроизведения - редупликации - генного вещества . Так возникла новая наука - молекулярная биология. Была сформулирована так называемая центральная догма молекулярной биологии: ДНК Ю РНК Ю белок. Смысл ее состоит в том, что генетическая информация, записанная в ДНК, реализуется в виде белков, но не непосредственно, а через посредство родственного полимера - рибонуклеиновую кислоту (РНК), и этот путь от нуклеиновых кислот к белкам необратим. Таким образом, ДНК синтезируется на ДНК, обеспечивая собственную редупликацию, то есть воспроизведение исходного генетического материала в поколениях; РНК синтезируется на ДНК, в результате чего происходит переписывание, или транскрипция, генетической информации в форму многочисленных копий РНК; молекулы РНК служат матрицами для синтеза белков - генетическая информация транслируется в форму полипептидных цепей. В специальных случаях РНК может переписываться в форму ДНК ("обратная транскрипция"), а также копироваться в виде РНК (репликация), но белок никогда не может быть матрицей для нуклеиновых кислот (подробнее см. ).
Итак, именно ДНК определяет наследственность организмов, то есть воспроизводящийся в поколениях набор белков и связанных с ними признаков. Биосинтез белка является центральным процессом живой материи, а нуклеиновые кислоты обеспечивают его, с одной стороны, программой, определяющей весь набор и специфику синтезируемых белков, а с другой - механизмом точного воспроизведения этой программы в поколениях. Следовательно, происхождение жизни в ее современной клеточной форме сводится к возникновению механизма наследуемого биосинтеза белков.
БИОСИНТЕЗ БЕЛКОВ
Центральная догма молекулярной биологии постулирует лишь путь передачи генетической информации от нуклеиновых кислот к белкам и, следовательно, к свойствам и признакам живого организма. Изучение механизмов реализации этого пути на протяжении десятилетий, последовавших за формулировкой центральной догмы, вскрыло гораздо более разнообразные функции РНК, чем быть только переносчиком информации от генов (ДНК) к белкам и служить матрицей для синтеза белков.
На рис. 1 представлена общая схема биосинтеза белка в клетке.
РНК-посредник
(messenger RNA, матричная РНК, мРНК), кодирующая белки, о
которой и шла речь выше, - это лишь один из трех главных классов клеточных
РНК. Основную их массу (около 80%) составляет другой класс РНК - рибосомные
РНК,
которые образуют структурный каркас и функциональные центры
универсальных белок-синтезирующих частиц - рибосом. Именно рибосомные РНК
ответственны - как в структурном, так и в функциональном отношении - за
формирование ультрамикроскопических молекулярных машин, называемых рибосомами.
Рибосомы воспринимают генетическую информацию в виде молекул мРНК и, будучи
запрограммированы последними, делают белки в точном соответствии с данной
программой.
Однако, чтобы синтезировать белки, одной только информации или программы недостаточно - нужен еще и материал, из которого их можно делать. Поток материала для синтеза белков идет в рибосомы через посредство третьего класса клеточных РНК - РНК-переносчиков (transfer RNA, транспортные РНК, тРНК). Они ковалентно связывают - акцептируют - аминокислоты, которые служат строительным материалом для беЛков, и в виде аминоацил-тРНК поступают в рибосомы. В рибосомах аминоацил-тРНК взаимодействуют с кодонами - трехнуклеотидными комбинациями - мРНК, в результате чего и происходит декодирование кодонов в процессе трансляции.
РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ
Итак, перед нами набор главных клеточных РНК, определяющих основной процесс современной живой материи - биосинтез белка. Это мРНК, рибосомные РНК и тРНК. РНК синтезируются на ДНК с помощью ферментов - РНК-полимераз, осуществляющих транскрипцию - переписывание определенных участков (линейных отрезков) двутяжевой ДНК в форму однотяжевой РНК. Участки ДНК, кодирующие клеточные белки, переписываются в виде мРНК, тогда как для синтеза многочисленных копий рибосомной РНК и тРНК имеются специальные участки клеточного генома, с которых идет интенсивное переписывание без последующей трансляции в белки.
Химическая структура РНК.
Химически РНК очень похожа на ДНК. Оба вещества - это линейные полимеры
нуклеотидов. Каждый мономер - нуклеотид - представляет собой фосфорилированный
N-гликозид, построенный из остатка пятиуглеродного сахара - пентозы, несущего
фосфатную группу на гидроксильной группе пятого углеродного атома
(сложноэфирная связь) и азотистое основание при первом углеродном атоме
(N-гликозидная связь). Главное химическое различие между ДНК и РНК состоит в
том, что сахарный остаток мономера РНК - это рибоза, а мономера ДНК -
дезоксирибоза, являющаяся производным рибозы, в котором отсутствует
гидроксильная группа при втором углеродном атоме (рис. 2).
 Рис. 2. Химические формулы остатков одного из рибонуклеотидов - уридиловой кислоты (U) и гомологичного ему дезоксирибонуклеотида - тимидиловой кислоты (dT) |
Азотистых оснований и в ДНК, и в РНК четыре вида: два пуриновых - аденин (А) и гуанин (G) -и два пиримидиновых - цитозин (С) и урацил (U) или его метилированное производное тимин (Т). Урацил характерен для мономеров РНК, а тимин - для мономеров ДНК, и это второе различие РНК и ДНК. Мономеры - рибонуклеотиды РНК или дезоксирибонуклеотиды ДНК - образуют полимерную цепь посредством формирования фосфодиэфирных мостиков между сахарными остатками (между пятым и третьим атомами углерода пентозы). Таким образом, полимерная цепь нуклеиновой кислоты - ДНК или РНК - может быть представлена как линейный сахаро-фосфатный остов с азотистыми основаниями в качестве боковых групп. Макромолекулярная структура РНК. Принципиальное макроструктурное различие двух типов нуклеиновых кислот состоит в том, что ДНК - это единая двойная спираль, то есть макромолекула из двух комплементарно связанных полимерных тяжей, спирально закрученных вокруг общей оси (см. [ , ]), а РНК - однотяжевой полимер. В то же время взаимодействия боковых групп - азотистых оснований - друг с другом, а также с фосфатами и гидроксилами сахаро-фосфатного остова приводят к тому, что однотяжевой полимер РНК сворачивается на себя и скручивается в компактную структуру , подобно сворачиванию полипептидной цепи белка в компактную глобулу. Таким способом уникальные нуклеотидные последовательности РНК могут формировать уникальные пространственные структуры. |
Впервые специфическая пространственная структура РНК была продемонстрирована при расшифровке атомной структуры одной из тРНК в 1974 г. [ , ] (рис. 3). Сворачивание полимерной цепи тРНК, состоящей из 76 нуклеотидных мономеров, приводит к формированию очень компактного глобулярного ядра, из которого под прямым углом торчат два выступа. Они представляют собой короткие двойные спирали по типу ДНК, но организованные за счет взаимодействия участков одной и той же цепи РНК. Один из выступов является акцептором аминокислоты и участвует в синтезе полипептидной цепи белка на рибосоме, а другой предназначен для комплементарного взаимодействия с кодирующим триплетом (кодоном) мРНК в той же рибосоме. Только такая структура способна специфически взаимодействовать с белком-ферментом, навешивающим аминокислоту на тРНК, и с рибосомой в процессе трансляции, то есть специфически "узнаваться" ими.

Рис. 3. Атомная (слева) и скелетная (справа) модели фенилаланиновой тРНК дрожжей
Изучение изолированных рибосомных РНК дало следующий разительный пример формирования компактных специфических структур из еще более длинных линейных полимеров этого типа. Рибосома состоит из двух неравных частей - большой и малой рибосомных субчастиц (субъединиц). Каждая субчастица построена из одной высокополимерной РНК и целого ряда разнообразных рибосомных белков. Длина цепей рибосомных РНК весьма значительна: так, РНК малой субчастицы бактериальной рибосомы содержит более 1500 нуклеотидов, а РНК большой субчастицы - около 3000 нуклеотидов. У млекопитающих, включая человека, эти РНК еще больше - около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно.
Было показано, что изолированные рибосомные РНК, отделенные от их белковых
партнеров и полученные в чистом виде, сами способны спонтанно сворачиваться в
компактные структуры, по своим размерам и форме похожие на рибосомные
субчастицы ].
Форма большой и малой субчастиц разная, и соответственно различается форма
большой и малой рибосомных РНК (рис. 4). Таким образом, линейные цепи
рибосомной РНК самоорганизуются в специфические пространственные структуры,
определяющие размеры, форму и, по-видимому, внутреннее устройство рибосомных
субчастиц, а следовательно, и всей рибосомы.
Минорные РНК. По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" .
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Рибозимы. Вся активная жизнь построена на обмене веществ - метаболизме, и все биохимические реакции метаболизма происходят с надлежащими для обеспечения жизни скоростями только благодаря высокоэффективным специфическим катализаторам, созданным эволюцией. На протяжении многих десятилетий биохимики были уверены, что биологический катализ всегда и всюду осуществляется белками, называемыми ферментами , или энзимами. И вот в 1982-1983 гг. было показано, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью [ , ]. Такие РНК-катализаторы были названы рибозимами. Представлению об исключительности белков в катализе биохимических реакций пришел конец.
В настоящее время рибосому тоже принято рассматривать как рибозим. Действительно, все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками. Идентифицирован каталитический участок большой рибосомной РНК, ответственный за катализ реакции транспептидации, посредством которой осуществляется наращивание полипептидной цепи белка в процессе трансляции.
Что касается репликации вирусных ДНК, то ее механизм мало чем отличается от редупликации генетического материала - ДНК - самой клетки. В случае же вирусных РНК реализуются процессы, которые подавлены или вовсе отсутствуют в нормальных клетках, где вся РНК синтезируется только на ДНК как на матрице. При инфекции РНК-содержащими вирусами ситуация может быть двоякой. В одних случаях на вирусной РНК как на матрице синтезируется ДНК ("обратная транскрипция"), а уж на этой ДНК транскрибируются многочисленные копии вирусной РНК. В других, наиболее интересных для нас случаях на вирусной РНК синтезируется комплементарная цепь РНК, которая и служит матрицей для синтеза - репликации - новых копий вирусной РНК. Таким образом при инфекции РНК-содержащими вирусами реализуется принципиальная способность РНК детерминировать воспроизведение своей собственной структуры, как это имеет место у ДНК.
Мультифункциональность РНК. Суммирование и обзор знаний о функциях РНК позволяют говорить о необыкновенной многофункциональности этого полимера в живой природе. Можно дать следующий список основных известных функций РНК.
Генетическая репликативная функция: структурная возможность копирования (репликации) линейных последовательностей нуклеотидов через комплементарные последовательности. Функция реализуется при вирусных инфекциях и аналогична главной функции ДНК в жизнедеятельности клеточных организмов - редупликации генетического материала.В целом РНК предстает перед нами столь удивительным полимером, что, казалось бы, ни времени эволюции Вселенной, ни интеллекта Творца не должно было бы хватить на ее изобретение. Как можно было видеть, РНК способна выполнять функции обоих принципиально важных для жизни полимеров - ДНК и белков. Неудивительно, что перед наукой и встал вопрос: а не могло ли возникновение и самодостаточное существование мира РНК предшествовать появлению жизни в ее современной ДНК-белковой форме?Кодирующая функция: программирование белкового синтеза линейными последовательностями нуклеотидов. Это та же функция, что и у ДНК. И в ДНК, и в РНК одни и те же триплеты нуклеотидов кодируют 20 аминокислот белков, и последовательность триплетов в цепи нуклеиновой кислоты есть программа для последовательной расстановки 20 видов аминокислот в полипептидной цепи белка.
Структурообразующая функция: формирование уникальных трехмерных структур. Компактно свернутые молекулы малых РНК принципиально подобны трехмерным структурам глобулярных белков, а более длинные молекулы РНК могут образовывать и более крупные биологические частицы или их ядра.
Функция узнавания: высокоспецифические пространственные взаимодействия с другими макромолекулами (в том числе белками и другими РНК) и с малыми лигандами. Эта функция, пожалуй, главная у белков. Она основана на способности полимера сворачиваться уникальным образом и формировать специфические трехмерные структуры. Функция узнавания является базой специфического катализа.
Каталитическая функция: специфический катализ химических реакций рибозимами. Данная функция аналогична энзиматической функции белков-ферментов.
ПРОИСХОЖДЕНИЕ ЖИЗНИ
Белково-коацерватная теория Опарина. Пожалуй, первая научная, хорошо продуманная теория происхождения жизни абиогенным путем была предложена биохимиком А.И. Опариным еще в 20-х годах прошлого века [ , ]. Теория базировалась на представлении, что все начиналось с белков, и на возможности в определенных условиях спонтанного химического синтеза мономеров белков - аминокислот - и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Основным ее постулатом было то, что спонтанно возникавшие в первичном "бульоне" белковоподобные соединения объединялись" в коацерватные капли - обособленные коллоидные системы (золи), плавающие в более разбавленном водном растворе. Это давало главную предпосылку возникновения организмов - обособление некой биохимической системы от окружающей среды, ее компартментализацию. Так как некоторые белковоподобные соединения коацерватных капель могли обладать каталитической активностью, то появлялась возможность прохождения биохимических реакций синтеза внутри капель - возникало подобие ассимиляции, а значит, роста коацервата с последующим его распадом на части - размножением. Ассимилирующий, растущий и размножающийся делением коацерват рассматривался как прообраз живой клетки (рис. 5).

Рис. 5.
Схематическое представление пути
происхождения жизни
согласно белково-коацерватной теории А.И.
Опарина
Все было хорошо продумано и научно обосновано в теории, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путем случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения - внутри коацервата и в поколениях - единичных, случайно появившихся эффективных белковых структур.
Мир РНК как предшественник современной жизни. Накопление знаний о генетическом коде, нуклеиновых кислотах и биосинтезе белков привело к утверждению принципиально новой идеи о ТОМ, что все начиналось вовсе не с белков, а с РНК [ - ]. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых, благодаря принципу комплементарности при синтезе новых цепей (подробнее см. ), обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки, могут быть генетическим материалом, то есть воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
По ряду соображений именно РНК, а не ДНК, могла представлять собой первичный генетический материал.
Во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды - продукты модификации рибонуклеотидов (см. рис. 2).
Во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа рибонуклеозид-полифосфатов (АТФ и т.п.).
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК.
В-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций. Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение - как модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности
РНК в происхождении жизни получила сильнейший толчок к развитию, и была
сформулирована концепция самодостаточного мира РНК,
предшествовавшего
современной жизни [ ,
].
Возможная схема возникновения мира РНК представлена на рис. 6.
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в олигомеры и полимеры типа РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Недавно А.Б. Четверин с сотрудниками (Институт белка РАН) экспериментально показали, что по крайней мере некоторые полирибонуклеотиды (РНК) в обычной водной среде способны к спонтанной рекомбинации, то есть обмену отрезками цепи, путем транс-эстерификации . Обмен коротких отрезков цепи на длинные, должен приводить к удлинению полирибонуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение (сплайсинг) олигонуклеотидов на комплементарной цепи как на матрице [ , ], означало становление механизма репликации РНК. Репликация самих РНК-катализаторов (рибозимов) должна была повлечь за собой возникновение самореплицирующихся популяций РНК. Продуцируя свои копии, РНК размножались. Неизбежные ошибки в копировании (мутации) и рекомбинации в самореплицирующихся популяциях РНК создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК - это "самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы" .
Возникновение биосинтеза белка. Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки (см. рис. 6).
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза по-липептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представлю свою версию процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 7), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации , приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью , то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.
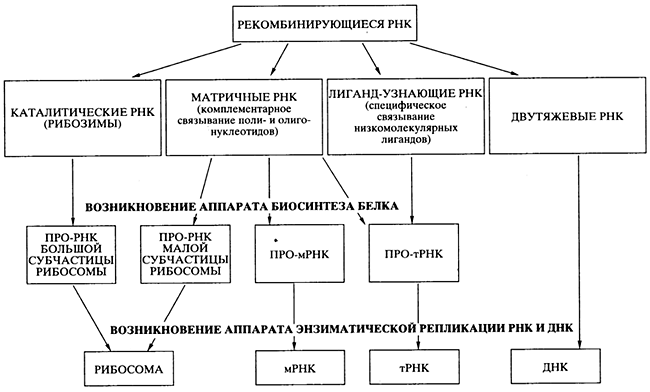
Рис. 7.
Схема эволюции и специализации молекул РНК
в процессе перехода от древнего мира РНК к современному миру
генетически детерминированного биосинтеза белков
Второй принципиальный момент в моей версии состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 7). Такая система уже могла синтезировать полипептидные цепи за счет катализируемой ею реакции транспептидации. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряженно, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов . Но это уже следующая сказка... ("И Шахразаду застигло утро, и она прекратила дозволенные речи" .)
Литература
1. Watson J.D., Crick F.H.C.
Molecular structure of
nucleic acids // Nature. 1953. V. 171. P. 738-740. 2. Watson J.D., Crick F.H.C.
Genetic implications of
the structure of deoxyribose nucleic acid // Nature 1953 V. 171. P. 964-967. 3. Спирин А.С.
Современная биология и биологическая
безопасность // Вестник РАН. 1997. № 7. 4. Spirin A.S.
On macromolecular structure of native
high-polymer ribonucleic acid in solution // Journal of Molecular Biology.
1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. et al.
Three-dimensional tertiary structure of yeast phenylalanine transfer RNA //
Science. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. et al.
Structure of yeast phenylalanine tRNA at 3 A resolution // Nature. 1974.
V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin
A.S.
Self-organization of ribosomal RNA // Sturcture, Function and
Genetics of Ribosomes / Eds. Hardesty B. and Kramer G. New York:
Springer-Verlag, 1986. P. 129-142. 8. Baserga SJ., Steitz J.A.
The diverse world of small
ribo-nucleoproteins // The RNA World / Eds. Gesteland R.F. and Atkins J.F. New
York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P.
359-381. 9. Kruger К., Grabowski PJ., Zaug AJ. et al.
Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA
intervening sequence of Tetrahymena
// Cell. 1982. V. 31. P. 147-157. 10. Guerrier-Takada С., Gardiner К., Marsh Т. et al.
The RNA moiety of ribonucleases P is the catalytic subunit of the enzyme
// Cell. 1983. V. 35. P. 849-857. 11. Опарин А.И.
Происхождение жизни. М.: Московский
рабочий, 1924. 12. Опарин А.И.
Возникновение жизни на Земле (3-е
изд.). М.: Изд-во АН СССР, 1957. 13. Woese С.
The evolution of the genetic code // The
Genetic Code. New York: Harper & Row, 1967. P. 179-195. 14. Crick F.H.C.
The origin of the genetic code //
Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E.
Evolution of the genetic apparatus //
Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W.
The RNA world // Nature. 1986. V 319
P. 618. 17. Joyce G.F., Orgel L.E.
Prospects for
understanding the origin of the RNA world // The RNA World / Eds. Gesteland
R.F. and Atkins J.F.
New York: Cold Spring Harbor Laboratory Press, Cold
Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I.,
Chetverin A.B.
Spontaneous rearrangements in RNA sequences // FEBS
Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W.
Isolation of new
ribozymes from a large pool of random sequences // Science. 1993. V. 261. P.
1411-1418. 20. Ekland E.H., Bartel D.P.
RNA-catalysed RNA
polymerization using nucleoside triphosphates // Nature. 1996 V. 382. P.
373-376. 21. Orgel L.E.
The origin of life - a review of facts
and speculations //Trends in Biochemical Sciences. 1998. V. 23. p. 491-495. 22. Альтштейн А.Д.
Происхождение генетической
системы: гипотеза прогенов // Молекулярная биология. 1987. Т. 21. С. 309-322.
Центральную догму биологии, сформулированную Фрэнсисом Криком в конце 1950-х, изучают в классическом виде: ДНК—>РНК—>белок. Но данных достаточно, чтобы усомниться в буквальном понимании этого главного принципа жизни.
Последний пример: в июньской публикации Scientific Reports российские ученые из Института биоорганической химии и Федерального научно-клинического центра физико-химической медицины показали, что разнообразие изоформ белков в клетках значительно меньше теоретически возможного. Журналисты поспешили сообщить, что меняется представление о центральной догме молекулярной биологии. Однако догма меняется уже 70 лет, потому что исходно была всего лишь гипотезой. Словом "догма" ее создатель Крик назвал потому, что нравилось это слово! Важно другое: как и почему меняется главная гипотеза молекулярной биологии.
Слишком много РНК
Генетическая информация считывается с кодирующих последовательностей генома, представленных генами. Только малая часть генома эукариот (растения, животные, грибы) содержит гены, а основная часть представлена протяженными нуклеотидными последовательностями с малоизученными функциями. В геноме человека только четверть покрыта генами и только 1% последовательностей ДНК кодирует информацию, записываемую в функциональных молекулах РНК (часть догмы "ДНК-->РНК"). То есть 1% геномной ДНК содержит информацию обо всех молекулах РНК. Зачем нужны оставшиеся 99%?
В последние годы стало понятно, что межгенные участки ДНК несут в себе регуляторную функцию: в них заложены системы и элементы, обеспечивающие тонкую настройку работы генов, их включение или выключение в определенных тканях организма или на конкретных стадиях развития. С такими элементами связываются различные комплексы, которые содержат молекулы регуляторных белков и РНК. Уже на этом уровне очевидно, что модель "ДНК-->РНК-->белок" полноценно не работает, так как основная масса ДНК не дает начала РНК, а несет другие функции.
Часть генов кодирует РНК с регуляторными функциями. Эти РНК не содержат информации о последовательности белка, а преимущественно организуют синтез белка в клетке. Основная часть таких РНК — компоненты рибосом (рибосомальные РНК), комплексы, осуществляющие трансляцию, а также молекулы-переносчики аминокислот (транспортные РНК), необходимые участникам процесса синтеза белка на матрице РНК (трансляции). 90% всей РНК клетки относится к перечисленным типам.
Среди оставшихся 10% молекул РНК представлены все белок-кодирующие РНК, но даже среди этих РНК найдены некодирующие молекулы, в частности, малые ядерные РНК. Эти РНК — необходимые компоненты комплекса сплайсинга. Сплайсинг — процесс удаления из первичной молекулы РНК некодирующих участков (интронов) и последовательного соединения кодирующих (экзонов); в итоге получается матричная РНК (мРНК), содержащая готовую к считыванию информацию о последовательности белка.
Именно этот комплекс готовит предшественников мРНК к синтезу правильных белков — путем вырезания из середины РНК последовательностей, не несущих в себе информации о составе белка, но содержащих регуляторные элементы. Так что и часть догмы "РНК --> белок" имеет свои ограничения.
Молекулярный "контроль качества"
А что мы знаем про так называемые "белок-кодирующие" гены? В клетках прокариот (бактерии) для такого типа генов все просто: на матрице ДНК транскрибируются молекулы РНК, на их базе идет синтез белковых молекул. Чаще всего молекулы РНК готовы для синтеза уже во время транскрипции.
В клетках эукариот все гораздо сложней: синтезирующиеся в процессе транскрипции молекулы РНК не готовы к трансляции (синтезу белка), прежде они должны претерпеть ряд изменений. Определенный набор модификаций вносится на концы молекул РНК (и РНК становится стабильной, а также попадает в определенные зоны клетки — "фабрики белка"), из середины молекул вырезаются интроны. Без сплайсинга и объединения экзонов правильную белковую молекулу не синтезировать.
С усложнением геномов вклад сплайсинга в процесс созревания мРНК увеличивается: у дрожжей только 4% белок-кодирующих генов подвергается сплайсингу, у дрозофилы — 83%, а у человека — 94%. Основная часть генов человека содержит более одного интрона в своем составе, и более половины генов человека могут сплайсироваться несколькими способами. Так что сплайсинг — это дополнительный регуляторный механизм, контролирующий количество "правильных" РНК, на матрице которых может запускаться синтез белковых молекул.
Кроме этого, сплайсинг часто является своего рода "контролем качества" молекул РНК, регулирует их стабильность. А так как альтернативный сплайсинг приводит к образованию на основе одной и той же молекулы РНК разных вариантов зрелых мРНК, это способ обеспечить дополнительное разнообразие белков в клетке. Такое разнообразие нужно для лучшей приспосабливаемости организма: разные изоформы белка могут работать в разных типах клеток, транспортироваться в разные компартменты или формировать разные поверхности узнавания для лигандов и т. д.
О чем "шумят" гены
Далеко не для всех изоформ белков известны функции, а во многих случаях для альтернативно сплайсированных молекул РНК не удается обнаружить белковый продукт. Авторы указанной статьи в Scientific Reports, изучавшие продукты альтернативного сплайсинга на модели мха, не обнаружили белков для большей части альтернативно сплайсированных молекул мРНК. В работах, выполненных на других модельных организмах, для многих альтернативно сплайсированных вариантов мРНК белковые молекулы также не были найдены.
Возможно, такие молекулы — побочный продукт регуляции "количества" генной экспрессии, "генный шум"; либо некоторые изоформы белка нужны в крайне ограниченных количествах.
Кроме этого, во многих интронах генов находятся регуляторные элементы, управляющие процессами сплайсинга, и там же могут находиться некодирующие РНК, участвующие в клеточном метаболизме. Так что и многообразие изоформ, и даже экспрессии белка могут контролироваться непосредственно молекулами РНК, без участия ДНК.
С развитием полногеномных технологий появляется все больше работ о некодирующих молекулах РНК. В геноме человека описан огромный пул таких РНК — "длинных" и "коротких": они осуществляют важные регуляторные функции в клетке. Эти РНК следят за стабильностью белок-кодирующих РНК, активируют или репрессируют гены, являются сенсорами при разных стрессах. Функции основной части некодирующих РНК еще не описаны, это целый мир, без которого клетка и организм не могут существовать.
Накопленные на сегодня данные наталкивают на мысль, что на молекулярном уровне жизнь — это форма реализации функций РНК. ДНК хранит информацию, белок отвечает за клеточный метаболизм, а жизнь клетки (и организма) организуется и контролируется на этапе функционирования молекул РНК.
Существуют даже предположения, что именно РНК на заре эволюции была первым биополимером, способным к самовоспроизводству. РНК, с одной стороны, подобно ДНК, способна быть хранилищем генетической информации (геномы огромной группы вирусов представлены РНК). С другой — известны и РНК с каталитической функцией, способные выполнять часть функций белков. Сторонники РНК-мира считают, что свойства РНК, позволившие им воспроизводить за счет собственной ферментной активности записанную в последовательностях нуклеотидов информацию, сыграли решающую роль в становлении генетического аппарата живых организмов.
Время для подобных обобщений еще не пришло. Ученые только начинают понимать, что система, которую они изучают уже 100 лет, гораздо сложней, чем казалось даже 20 лет назад.
Оксана Максименко, кандидат биологических наук, Институт биологии гена РАН
Если очень коротко, то это предположение, что жизнь на Земле начиналась не с ДНК или белков, а с ансамблей РНК.
Главной проблемой в происхождении жизни является вопрос о возникновении аппарата наследственности и вещества наследственности – нуклеиновых кислот.
Маловероятно, что жизнь началась с ДНК, потому что это громоздкие двухцепочечные молекулы, просто здоровенные по молекулярным меркам. Кроме того, они не могут сами себя обслуживать (нужны белки, а это всё усложняет).
Если предположить, что жизнь началась с белков, то возникают другие проблемы (например, откуда бы им взяться без ДНК).
Чем же РНК лучше?
РНК – одноцепочечные гибкие молекулы. Долгое время считалось, что функция РНК состоит в простом считывании информации с цепей ДНК + последующий синтез белков на РНК-матрице. Потом оказалось, что кодирующие РНК - малая доля всех РНК, а основная часть - как раз некодирующие. Было открыто явление обратной транскрипции – синтез ДНК на матрице РНК. Так же стало известно, что РНК способны самосворачиваться в компактные структуры, узнавать и избирательно связывать органические молекулы. Совсем как… белки.
Получается, РНК - уникальный биополимер, которому свойственны функции ДНК и белков одновременно :
1) Комплементарная репликация (если для чайников, то это достройка «обратной цепи» по имеющейся цепи. Не копирование и не зеркальное построение, а как бы «снятие негатива»)
2) Кодирование белков
3)Структурная и формообразующая функции (сворачивание в клубочки разной формы)
4)Специфическое узнавание молекул
5) Катализ реакций
Выходит, молекулы РНК могли бы обходиться не только без ДНК как генетического вещества, но и без белков для осуществления катализа реакций. Эта идея была сформулирована в 1986 г. и в настоящее время является почти общепринятой.
В древнем мире РНК не было ни белков, ни ДНК, а лишь ансамбли различных молекул РНК. Дело в том, что из единичных молекул РНК в подходящих условиях вырастают колонии из копий исходной молекулы.
Возможно, эти процессы происходили в мелких лужицах, которые то подсыхали и разделялись, то снова заливались доисторическим дождиком, сливаясь в одну большую. В луже, содержащей РНК, молекулы копировались (иногда с ошибками) и накапливались. Но на этом этапе никакой эволюции еще не было, потому что любые варианты копировались одинаково интенсивно. Чтобы естественный отбор начал работать, необходима была какая-то форма обособления отдельных ансамблей РНК. "Лучшие" обособленные ансамбли РНК должны расти быстрее других, перерастать их, тем самым обеспечивая отбор. Поэтому в большинстве теорий происхождения жизни возникновение каких-то ограничивающих мембран - необходимое условие начала эволюции. В этом одна из основных трудностей таких теорий.
Но для РНК была экспериментально показана способность формировать молекулярные колонии на гелях или других влажных твердых средах. Смешанные колонии РНК на твердых или полутвердых поверхностях, без каких-то ни было мембран, и могли быть первыми эволюционирующими бесклеточными ансамблями. Это создавало условия для очень быстрой эволюции: колонии РНК не были отгорожены от внешней среды и могли легко обмениваться своим генетическим материалом.
И так, эволюционный процесс должен был быть исключительно быстрым благодаря трем обстоятельствам :
1) Спонтанные перестройки молекул РНК + низкая точность копирования = огромное число вариантов для отбора.
2) Свободный обмен молекулами РНК между колониями через воду и атмосферу делал любые полезные инновации достоянием всех и позволял колониям быстро совершенствоваться за короткое время.
3) Экспоненциальное обогащение всей популяции «лучшими» молекулами РНК в циклах умножения и отбора - эволюционный двигатель для всего коммунального мира РНК в целом.
Если именно мир РНК возник и эволюционировал в клеточные формы жизни на Земле, то весь путь эволюции до индивидуальных организмов с клеточной структурой, ДНК и современным аппаратом белкового синтеза должен был быть пройден менее чем за пол-миллиарда лет.
Это адаптированное сокращение доклада А.С. Спирина "Древний мир РНК". Гуглится легко. Рекомендую прочитать его целиком, если вам интересны подробности.
Мир РНК: вчера и сегодня
Долгое время считали, что рибонуклеиновая кислота (РНК), ближайшая «родственница» знаменитой ДНК, выполняет в организме служебные функции, являясь лишь посредником в сложнейших внутриклеточных процессах. И хотя признавалось, что на ранних этапах эволюции жизни РНК могла играть главенствующую роль, но казалось очевидным, что она давно уступила первенство более специализированным молекулам – катализаторам и носителям информации. Однако открытие множества регуляторных РНК, связанных с феноменом некодирующего «темнового генома», буквально перевернуло представления о современном «мире РНК» и послужило толчком к поиску и созданию новых диагностических и лекарственных средств
Классический взгляд на процесс реализации наследственной информации в живой клетке сформировался еще в начале второй половины XX в. Согласно нему, вся наследственная информация, определяющая жизнь организма, закодирована в виде последовательности нуклеотидов в особом биополимере – дезоксирибонуклеиновой кислоте (ДНК). «Размножение» и передача этой информации происходит путем удвоения молекул ДНК с участием специальных белков-ферментов.
Когда участок ДНК, в котором содержится информация о строении того или иного белка, – ген – начинает «работать», с него с помощью специальных ферментов считывается «промежуточная» информация в виде молекул рибонуклеиновой кислоты (РНК). Нуклеотидная последовательность этой РНК служит «программой»-матрицей для особых молекулярных машин – рибосом , которые приступают к синтезу соответствующего белка
В этой схеме преобразования генетического кода в конечный «полезный продукт» центральным элементом является ДНК. Была даже сформулирована «центральная догма молекулярной биологии», гласившая, что поток информации в живой клетке идет по направлению от ДНК к РНК и далее к белку, и только в этом направлении: синтезировать ДНК или РНК, соответствующую определенному белку, клетка не умеет. Неудивительно, что увлекательная книга молекулярного биолога М. Д. Франк-Каменецкого «Самая главная молекула», вышедшая в 1983 г., была посвящена именно ДНК.
Важная роль отводилась также белкам, как главным строительным «кирпичикам» и, в первую очередь, катализаторам, способным в условиях живой клетки опосредовать самые разные биохимические реакции и процессы. РНК же представлялась скорее «рабочей» молекулой, расходным материалом белкового синтеза. Конечно, помимо матричной РНК были известны и транспортные РНК, переносящие аминокислоты на рибосомы, а также РНК в составе рибосомного каркаса, однако и им отводилась роль скромных «золушек».
Со временем стали накапливаться наблюдения, которые заставляли задуматься: а так ли просто все устроено в геноме? К примеру, оказалось, что у некоторых вирусов наследственная информация зашифрована не в ДНК, а в РНК, и они способны синтезировать ДНК по «матрице» РНК при встраивании своего генетического материала в геном клетки-хозяина. Однако несмотря на отдельные противоречия, взгляды на природу и роль важнейших биополимеров, вошедшие во все школьные учебники, десятилетиями оставались незыблемыми.
Между тем, невзирая на трудности экспериментов с РНК, молекулы которой менее стабильны, чем ДНК и белков, исследователи продолжали упорно изучать эти «немодные» биополимеры. И в конце XX в. классические научные представления начали рушиться под напором новых поразительных открытий.
Жизнь начиналась с РНК
После расшифровки генома человека и других высокоорганизованных животных оказалось, что большая часть ДНК не содержит генов, кодирующих белки. (К слову, когда сравнили геномы различных млекопитающих, оказалось, что по набору генов «царь природы» немногим отличается от той же мыши.) В геноме человека длиной около трех миллиардов пар нуклеотидов ожидали обнаружить более 100 тыс. генов, но их оказалось по крайней мере в три раза меньше! Причем в это число входят все гены, кодирующие белки, а также гены рибосомных и транспортных РНК. Вместе с известными на то время регуляторными последовательностями все эти гены занимают не более 1,5 % геномной ДНК. А что же остальная часть?
С полным пренебрежением к непонятному такую некодирующую ДНК – так называемый темновой геном – стали считать «мусором», накопившимся в процессе эволюции. И хотя впоследствии было обнаружено, что с участков «мусорной» ДНК все же считывается какая-то информация и синтезируется огромное количество РНК, это явление было оценено как «транскрипционный шум».
Самым удивительным открытием стал тот факт, что белки не являются монополистами в умении катализировать биологические реакции. Так, в ядре клетки обнаружился набор небольших РНК, которые при участии белков обеспечивают процесс созревания матричной РНК. Дело в том, что с гена считывается длинная РНК, которую необходимо нарезать на фрагменты и затем определенным образом «сшить», чтобы получилась нужная программа. Правильность этой «сшивки» и обеспечивают специальные РНК.

Был открыт и каталитически активный РНК-белковый комплекс теломераза , благодаря которому происходит синтез концевых участков хромосом. Оказалось, что в рибосоме РНК также выполняет не только структурную роль – из нее построен сам каталитический центр! Таким образом, все ключевые функции в процессе белкового синтеза принадлежат молекулам РНК.
Выяснилось, что РНК, подобно белкам, могут формировать компактные структуры, способные к высокоспецифичному взаимодействию практически с любыми маленькими и большими молекулами. РНК-катализаторы были найдены в природе, их используют как вирусы, так и клетки высших организмов. А вскоре самые различные каталитические РНК научились получать искусственным путем.
Факты упрямо указывали на первенство РНК как биологической молекулы. Да, ДНК является стабильным хранителем информации, но не может функционировать без помощи белков. Белки – эффективные катализаторы, но на роль носителей информации принципиально не годятся. А вот РНК оказалась именно такой универсальной, многофункциональной молекулой, какой и должна быть «прародительница».
Стало ясно, что именно из простых РНК-содержащих систем произошел современный биологический мир. Но куда же делся этот «мир РНК», когда на смену пришли более специализированные молекулы? Не являются ли сегодня РНК своего рода «молекулярными окаменелостями», имеющими хотя и важное, но ограниченное значение? Еще совсем недавно ответ на эти вопросы сомнений не вызывал.
Тридцать лет спустя
 В 1997 г. в Германии состоялась конференция, посвященная исследованиям РНК. Ей предстояло подвести итог масштабной программы исследований РНК: все, что касалось этих биополимеров, представлялось на тот момент ясным и понятным. В этой атмосфере настоящим «взрывом бомбы» прозвучал доклад об открытии РНК-интерференции
– ранее неизвестного механизма регуляции функционирования генов с помощью специальных малых РНК.
В 1997 г. в Германии состоялась конференция, посвященная исследованиям РНК. Ей предстояло подвести итог масштабной программы исследований РНК: все, что касалось этих биополимеров, представлялось на тот момент ясным и понятным. В этой атмосфере настоящим «взрывом бомбы» прозвучал доклад об открытии РНК-интерференции
– ранее неизвестного механизма регуляции функционирования генов с помощью специальных малых РНК.
Вообще-то удивительно, что обнаружение этого довольно очевидного механизма потребовало так много времени. Ведь теоретически было давно известно, что остановить работу целевой генетической программы, т. е. «заблокировать» конкретную нуклеиновую кислоту, можно путем присоединения к ней небольшого комплементарного фрагмента РНК или ДНК, достаточного по размеру, чтобы обеспечить избирательность действия.
Именно такой способ избирательного регулирования функций нуклеиновых кислот был впервые предложен еще в 1960-х гг. новосибирским исследователем Н. И. Гриневой и ее коллегами, работавшими под руководством будущего академика РАН Д. Г. Кнорре. Ученые справедливо решили, что лучший способ направленно воздействовать на гены – использовать так называемые антисмысловые олигонуклеотиды, т.е. короткие комплементарные фрагменты ДНК или РНК (Belikova et al. , 1967).
И вот десятилетия спустя было доказано, что этот механизм направленного воздействия на генетический материал функционирует в живых системах при регуляции экспрессии генов. Это тот редкий случай в истории науки, когда явление сначала было описано теоретически, затем его схема реализована в эксперименте и лишь десятилетия спустя оно обнаружено в природе.
Забегая вперед, нужно отметить, что своими работами сибирские ученые открыли новое направление прикладной молекулярной биологии, которое сегодня бурно развивается во всем мире. Уже получены первые фармацевтические препараты на основе интерферирующих микроРНК, а в будущем ожидается создание широчайшего спектра лекарств, в том числе и для избирательной инактивации уже самих микроРНК (такие ингибиторы даже получили специальное название – «анти-мир») и других некодирующих РНК.

На той же конференции 1997 г. было заявлено об обнаружении в клетках мозга и других органов множества разнообразных некодирующих РНК, концентрации которых меняются в зависимости от состояния организма. Стало очевидно, что речь должна идти не о свертывании, а о расширении дальнейших исследований РНК.
Сокровища «мусорной» ДНК
Очень скоро были получены данные о чрезвычайно широком распространении РНК-интерференции в природе и начаты работы по практическому использованию интерферирующих РНК для создания терапевтических препаратов и генетически модифицированных организмов.
Более того, было показано, что в организме синтезируется огромное число – тысячи видов – некодирующих РНК, информация о которых содержалась в той самой якобы ненужной «мусорной ДНК». Были обнаружены микроРНК-регуляторы активности генов, макроРНК, выполняющие различные регуляторные функции, а также множество других РНК с пока неизвестными функциями. Удивительно, что многие РНК находятся не только в клетках тканей, но и в крови, и способны «путешествовать» по организму.
Длинные некодирующие РНК пока что относительно мало изучены, и механизм их действия установлен лишь в некоторых случаях. Исследователи склоняются к заключению, что такие РНК играют роль организаторов сложных функциональных структур, включающих различные биомолекулы. Молекула РНК идеально подходит на подобную роль: она может содержать в себе различные модули, которые способны связываться как с конкретными белками, так и с участками ДНК. Такие РНК-модули могут располагаться определенным образом относительно друг друга, благодаря чему появляется возможность создавать супрамолекулярные ансамбли любой степени сложности. Самый наглядный пример такой структуры – рибосома.
Таким образом, именно благодаря РНК создаются основные структуры клетки, делающие ее «живой». Длинные некодирующие РНК управляют также работой генетического аппарата клетки. Они включают и выключают работу целых областей хромосомы, определяя точки специфической модификации хромосомных белков.


С состоянием организма тесно связаны и концентрации разнообразных микроРНК в органах и тканях. Эти РНК, участвуя в регуляции активности огромного числа генов, воздействуют на важнейшие физиологические и метаболические процессы, протекающие в отдельной клетке и в организме в целом.
Например, с помощью определенного набора микроРНК можно изменить степень клеточной дифференциации – получить из специализированных клеток недифференцированные стволовые либо, напротив, управлять дифференциацией стволовых клеток в нужном направлении. Доказано, что с помощью микроРНК регулируется дифференциация клеток жировой ткани, метаболизм липидов, секреция инсулина и других гормонов и т. д.
Судя по последним данным, в сферу «юрисдикции» РНК попадает свыше 60 % всех генов. Фактически, именно микроРНК «руководят» развитием целых органов и организма в целом, запуская одни процессы и останавливая другие по соответствующим «биологическим часам».
В составе специальных транспортных мембранных структур, таких как экзосомы , микроРНК могут передаваться не только между тканями и органами одного организма, но даже между организмами. Например, в молоке матери содержатся экзосомы с микроРНК, которые играют важную роль в становлении иммунной системы ребенка, причем уровень их секреции особенно высок в первые шесть месяцев лактации (Kosaka et al. , 2010).

Неудивительно, что интерес к таким транспортным структурам и составу переносимых ими микро- и других некодирующих РНК сегодня крайне высок, и недавно начатые исследования были подхвачены тысячами ученых из разных стран. Было даже организовано международное общество по внеклеточным везикулам, а в апреле 2012 г. в Швеции состоялась первая конференция по этой тематике, где было объявлено о создании специализированного журнала.
Точки приложения
Итак, некодирующие РНК являются главными регуляторами, управляющими функциями генов и целых генетических ансамблей, при этом число известных некодирующих РНК постоянно растет. На их концентрацию и соотношение влияют и протекающие в организме патологические процессы (например, опухолевые). Отсюда очевидна перспективность применения методов количественного анализа некодирующей РНК для целей медицинской диагностики. А подавление активности РНК, ассоциированной с развитием болезни, должно стать новым подходом в терапии.
Сегодня известно, что с помощью определенных микроРНК можно блокировать развитие рака предстательной и молочной желез (Tavazoie et al. , 2008). МикроРНК рассматриваются и в качестве потенциальных лечебных средств в терапии сердечно-сосудистых заболеваний (Latronico & Condorelli, 2009). Ведутся клинические исследования экзосом, содержащих микроРНК, которые циркулируют в крови при меланомах и различных формах рака легких (Rabinowits et al. , 2009).
Подобные исследования активно ведутся и в Сибирском отделении РАН. Благодаря разработанной в Институте химической биологии и фундаментальной медицины СО РАН оригинальной методике высокопроизводительного секвенирования РНК появилась возможность быстро и точно получать информацию о профиле микроРНК в различных клинических образцах. Уже опубликованы данные о спектрах микроРНК здоровых людей (Semenov et al. , 2012), ведется работа по определению соответствующих РНК-профилей при различных патологиях.

В ИХБФМ получены и обнадеживающие экспериментальные данные о возможности проведения противоопухолевой и антиметастатической терапии ферментом РНКазой А, который влияет на концентрацию и спектр опухолевых микроРНК в клетках и сыворотке крови больных животных (Mironova et al. , 2012).
Исследованиями микроРНК с целью развития диагностических методов занимаются и в недавно организованном Институте молекулярной и клеточной биологии СО РАН (Новосибирск). Например, там был изучен профиль экспрессии некоторых микроРНК при различных опухолях щитовидной железы.
Детальное изучение свойств РНК подтолкнуло исследователей к созданию новых технологий, обещающих принести огромную пользу человечеству уже в ближайшем будущем. Речь идет о широко известной технологии SELEX, позволяющей получать молекулы РНК с заданными свойствами – аптамеры , которые способны селективно и прочно связываться с молекулой-мишенью. Суть аптамерной технологии в том, что сначала химико-ферментативными методами создаются так называемые РНК-библиотеки случайных последовательностей, содержащие огромное множество разнообразных молекул РНК. Затем из этого множества методами молекулярной селекции по способности специфично взаимодействовать с целевой молекулой отбирают аптамеры и нарабатывают их в нужном количестве, хотя искомая РНК может быть представлена в составе библиотеки единственной молекулой.
Сегодня в качестве аптамеров и терапевтических средств используется не только РНК с обычной «природной» структурой: с помощью различных химических модификаций можно получить молекулы, устойчивые к действию разрушающих РНК ферментов, которых много в биологических средах. Такие «искусственные» РНК имеют большие перспективы в диагностике заболеваний: например, можно получить РНК-аптамеры к молекулам – маркерам заболеваний и на их основе создать биосенсоры, обладающие уникальной чувствительностью.

Сочетание основанных на РНК биоаналитических подходов к диагностике заболеваний с новыми -терапевтическими средствами на основе микроРНК и некодирующих РНК в ближайшем будущем должно привести к новому прорыву в медицине.
С каждым годом исследователи открывают все новые и новые грани удивительного «мира РНК». Высокопроизводительные технологии секвенирования нуклеиновых кислот позволили обнаружить в клетках и крови огромное множество разнообразных РНК, среди которых оказались транскрипты тех участков ДНК, которые ранее считались «молчащими».
В результате в конце XX в. удалось открыть природный механизм огромной значимости, управляющий работой клеточного генома, который задолго до этого события был предвосхищен сибирскими учеными в идее «антисмысловых нуклеотидов».
Раскрытие многих тайн «мира РНК», который долгое время скрывался от исследователей за ширмой более узкоспециализированных молекул, белков и ДНК, приблизило нас к пониманию фундаментальных вопросов происхождения жизни. И более того: этот мир РНК, претворившийся в новые перспективные биомедицинские технологии, постепенно становится частью повседневной жизни человечества.
Литература
Власов В. В., Власов А. В. Жизнь начиналась с РНК // НАУКА из первых рук. 2004. № 2 (3). C. 6-19.
Власов А. В. Эволюция в пробирке // НАУКА из первых рук. 2006. № 1 (7). С. 50-59.
Власов В. В. Лекарство для генов // НАУКА из первых рук. 2007. № 2 (14). С. 56-59.
Карпова Г. Г., Грайфер Д. М., Малыгин А. А. Рибосома – минифабрика по производству белков // НАУКА из первых рук. 2006. № 6 (12). С. 46-53.
Франк-Каменецкий М. Д. Самая главная молекула, Москва: Наука, 1983.
Черноловская Е. Л. РНК-интерференция: клин клином… // НАУКА из первых рук. 2008. № 1 (19). С. 54-59.
Knorre D. G., Vlassov V. V., Zarytova V. F. et al. Design and targeted reactions of oligonucleotide derivatives, Boca Raton, CRC Press, 1994.
Vlassov V. V., Pyshnyi D. V., Vorobjev P. E. Nucleic acids: structures, functions, and applications. In Handbook of nucleic acids purification, Ed. D. Liu, Boca Raton, CRC Press, 2009.